
第一作者:申创
通讯作者:辛俊亮
通讯单位:香港正版资料图库1
论文链接:https://doi.org/10.1016/j.jhazmat.2023.133365
文章亮点:
•高镉辣椒品种通过提高Nramp5的表达增加了根系吸收镉的能力。
•高镉辣椒品种通过氮和激素代谢途径消除镉毒性。
•低镉辣椒品种通过根细胞壁木质素合成相关基因来减少镉积累。
•低镉辣椒品种通过硫代谢,液泡区隔和胞外排出来减少镉毒性。
•根细胞壁木质素合成相关基因CaCOMT1在拟南芥中表达减少了镉积累。
研究内容:
辣椒品种间镉积累差异的分子机制尚不清楚。本研究对高镉(J)和低镉品种(Z)在镉浓度分别为0和0.4 mg/L的条件下进行了根系转录组比较分析。结果表明,高镉品种通过提高Nramp5的表达来促进根对镉的吸收,并通过增加NIR1、GLN1和IAA9等基因的表达来消除镉毒性。低镉品种通过增强细胞壁木质素合成基因PAL、COMT、4CL、LAC和POD以及镉转运蛋白ABC、MTP1和DTX1来减少镉积累。在拟南芥中,硫代谢相关基因的表达升高,可能有助于其解毒镉的能力。为了调查CaCOMT1的功能,我们构建了一个拟南芥过表达系(OE-CaCOMT1)。结果表明, OE-CaCOMT1将木质素含量显著提高了38–42%,镉向地上部分转运减少了32%。本研究通过转录组分析为辣椒镉积累机制提供了全面的了解。此外,本研究阐明了CaCOMT1的关键功能,为低镉蔬菜的安全生产提供理论依据(图1)。
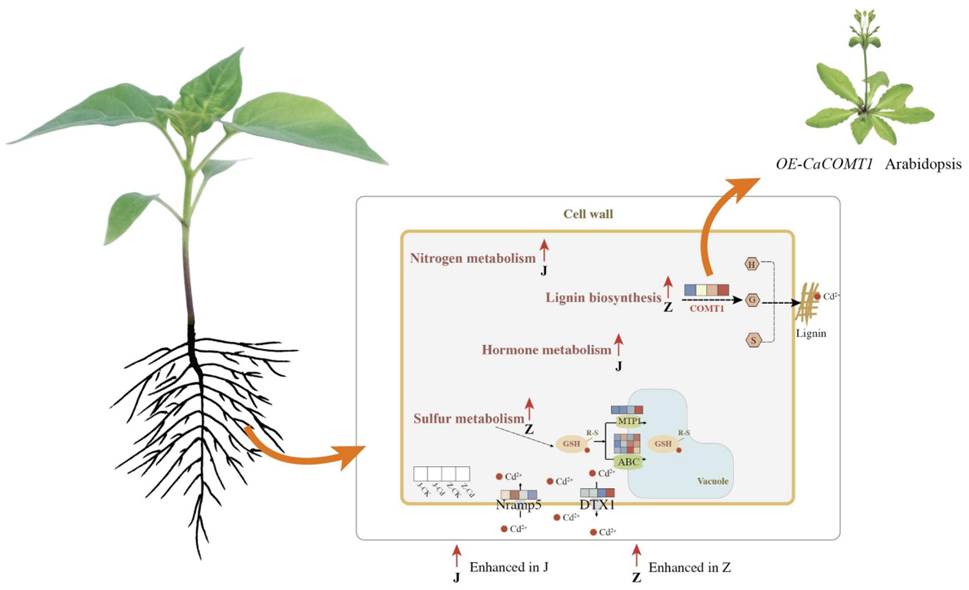
图1 图文摘要
转运蛋白相关基因的表达通过影响镉的分布和运输来影响植物对镉的耐受性和积累。差异表达基因分析显示,位于细胞质膜上负责吸收镉的Nramp5在J-Cd中的表达高于Z-Cd。与对照相比,镉处理下J和Z中定位于液泡膜上负责Cd输入的转运蛋白,如ABC转运蛋白(ABCC1、ABCC2和ABCC4)和金属耐受蛋白(MTP1)在镉处理下均高表达(图2)。此外,镉胁迫下,ABCs和MTP1在Z中的表达显著高于J (图2)。DTX1是解毒外排载体(DTX)家族的成员,位于细胞膜上,负责镉外排,无论植物是在对照条件下还是镉处理下,其在Z中的表达水平显著高于J(图2)。这些结果表明,镉吸收和外排转运蛋白的表达差异,以及液泡隔离,是辣椒品种间镉积累差异形成的关键因素。
细胞壁木质素合成相关基因苯丙氨酸氨解基因(PALA和PAL5)、咖啡酸O-甲基转移酶1 (COMT1)、4-香豆酸酯:辅酶A连接酶(4CL1、4CL2和4CL5)、肉桂酰辅酶A还原酶(CCR1和CCR2)、过氧化物酶(POD12、POD52和POD72)和漆酶(LAC3和LAC7)在Z中的表达量均显著高于J(图2)。镉胁迫下,与细胞壁果胶合成和果胶甲酯酶(PME1、PME7和PME51)相关的基因在Z中的表达量显著高于J(图2)。此外,木葡聚糖内糖基化酶/水解酶(XTH9和XTH30)、可以松弛细胞壁结构,促进细胞壁合成的基因在Z中的表达量明显高于J(图2)。这些发现说明,Z通过细胞壁合成和增厚来应对镉胁迫,而J主要通过果胶甲酯酶来调节细胞壁吸附镉的能力。

图2 对照(CK)和镉处理下(Cd)辣椒品种间差异表达的基因
(注:表达热图从左至右依次为J-CK、J-Cd、Z-CK、Z-Cd。每个差异表达基因的表达水平是三个生物样本的平均值,然后进行z-score标准化。)
对照条件下,OE-CaCOMT1和野生型拟南芥的生长情况相似,茎长和根长差异不显著 (图3A、3C、3D)。但OE-CaCOMT1拟南芥的生物量显著高于野生型拟南芥 (图3B)。镉胁迫下,OE-CaCOMT1和野生型拟南芥的生物量分别较对照条件下显著降低23.9%和27.9% (图3B)。镉处理下OE-CaCOMT1拟南芥的茎长和根长与对照条件下无显著差异 (图3C和3D)。相比之下,野生型植株的茎长和根长分别比对照条件下降低了7.3%和12.4% (图3C和3D)。这些结果表明,CaCOMT1增强了拟南芥对镉胁迫的耐受性。
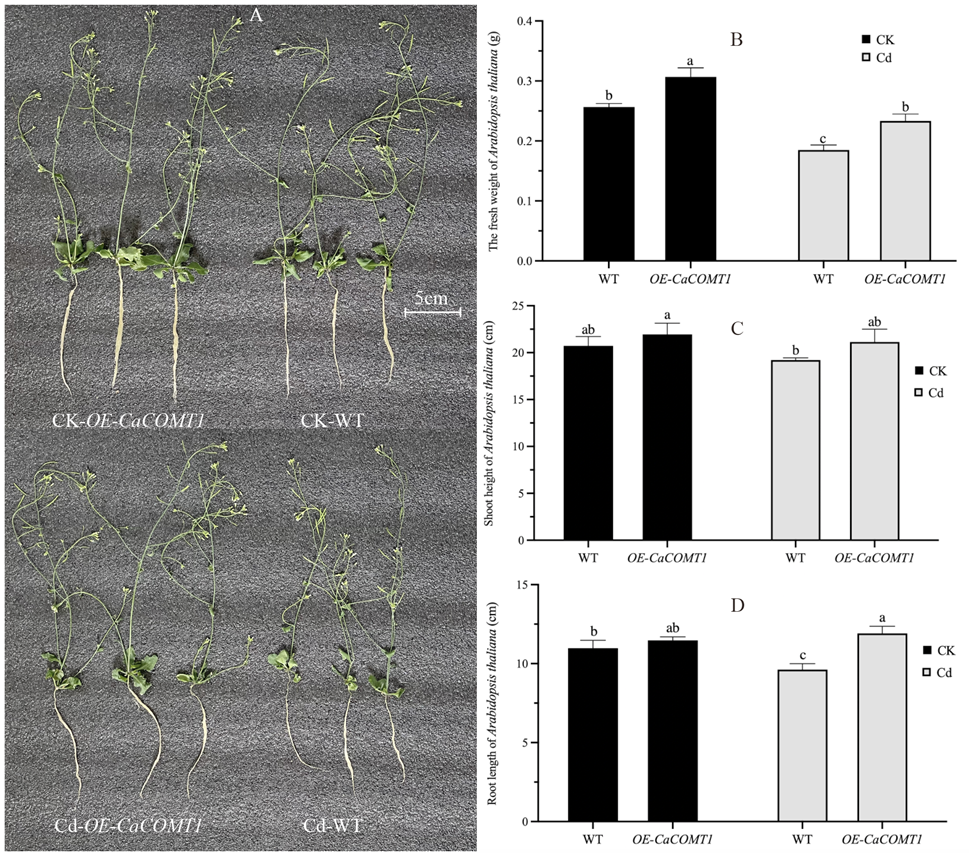
图3 对照和镉处理下,拟南芥的生长情况(A)、鲜重(B)、茎长(C)和根长(D)。
(注:数值表示平均值±标准差(n = 3)。不同小写字母表示处理间差异在p<0.05水平上显著。)
OE-CaCOMT1拟南芥细胞壁木质素的总体吸光值高于野生型拟南芥(图4A)。在1328 cm-1和1125 cm-1处,分别代表丁香基(S)单体芳环上的C-O和C-H基团的吸光值在OE-CaCOMT1 拟南芥中比在野生型拟南芥中高。此外,在1265 cm-1处,代表愈创木基的芳香环(G)单体的吸光值在OE-CaCOMT1 拟南芥中也比在野生型拟南芥中高(图4A)。上述结果表明,CaCOMT1过表达可增加拟南芥根细胞壁中木质素S和G单体的含量。
在对照条件下,野生型拟南芥中检测不到CaCOMT1的表达,只有AtCOMT1可以被检测到(图4B)。对照下,在OE-CaCOMT1拟南芥中CaCOMT1的表达量比AtCOMT1高80倍(图4B)。此外,在OE-CaCOMT1拟南芥中,AtCOMT1的表达量比野生型拟南芥显著降低32%(图4B),说明CaCOMT1可以抑制拟南芥中AtCOMT1的表达。镉处理显著提高了野生型和OE-CaCOMT1拟南芥的AtCOMT1,但CaCOMT1的表达不受影响(图4B)。
对照下,野生型和和OE-CaCOMT1拟南芥的木质素含量分别为142.91和204.14 mg/g (干重),镉处理下分别为176.25和243.66 mg/g (干重)(图4C)。此外,根组织化学染色显示OE-CaCOMT1拟南芥木质部和髓细胞壁木质素沉积明显增加,颜色呈深粉红色(图4D)。这些结果与木质素含量一致,说明拟南芥木质素含量与COMT1表达呈正相关。
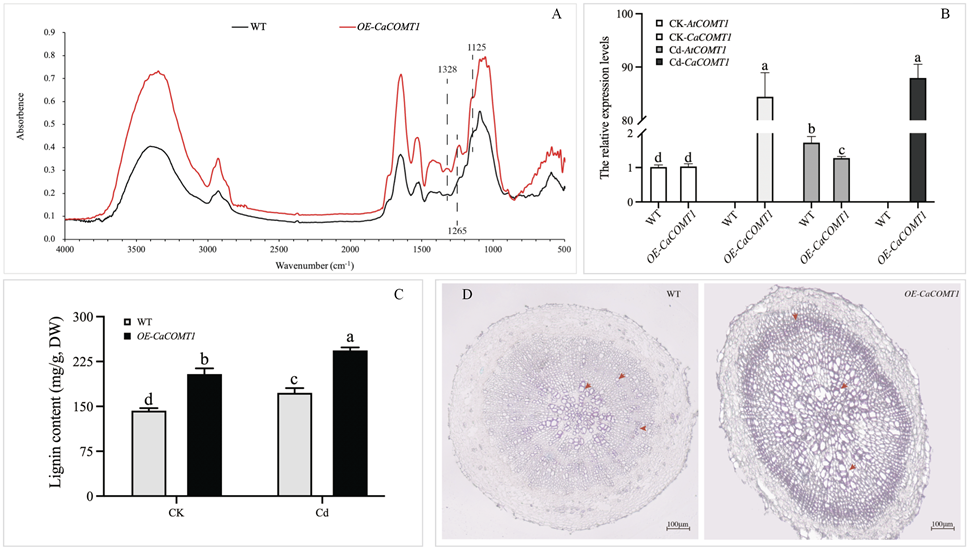
图4 对照和镉处理下,野生型和OE-CaCOMT1拟南芥根细胞壁红外光谱分析(A)、AtCOMT1和CaCOMT1的表达水平(B)、木质素含量(C)和组织化学染色(D)
本研究发现辣椒高镉品种主要通过提高Nramp5的表达水平来促进根吸收镉,这可能是其高镉积累的重要原因。此外,高镉品种在体内对镉毒性的反应是通过提高参与氮代谢、硫代谢和植物激素途径(包括生长素和脱落酸)的基因表达来实现的。低镉品种主要通过增强细胞壁木素合成、抗氧化酶、转运蛋白外排和液泡区隔来减少镉进入根细胞,这也与低镉品种低积累镉密切相关。此外,从木质素合成途径中筛选出的关键基因CaCOMT1在拟南芥中过表达后显示,CaCOMT1显著提高了镉胁迫下拟南芥的生物量和木质素含量。此外,CaCOMT1在辣椒的镉固定和转运中起着重要作用。总之,本研究揭示了CaCOMT1在镉固定中的潜在作用,建议开展更广泛和更深入的研究,以充分了解通过转录组分析鉴定的各种基因在植物吸收和固定重金属中的明确机制。本研究结果为低镉积累蔬菜品种的筛选和遗传改良提供了潜在的切入点。